How to Track the Artemis II Mission
And eavesdrop on mission control

Are Gossiping Mushrooms Sharing Your Public Urination Secrets?
Either way, show a little decorum, okay?

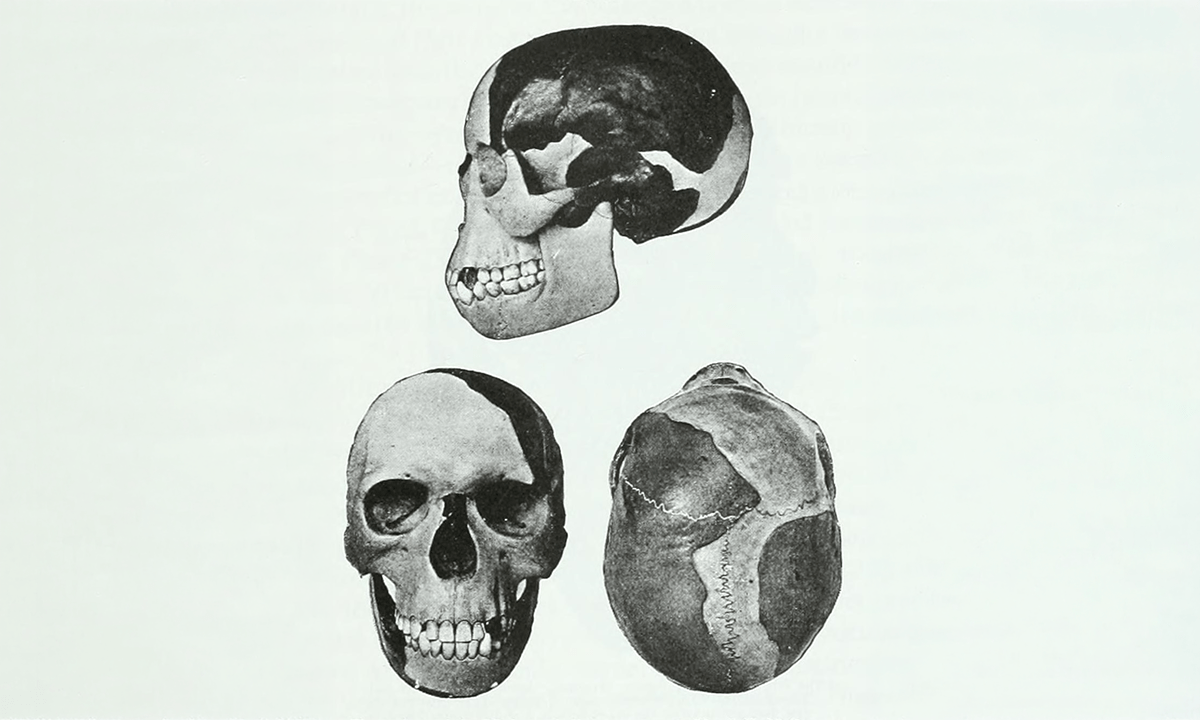
When Dogs First Became Man’s Best Friend
Ancient canid DNA pushes date of dog domestication back millennia

Meet the Arthropod That Originated Fangs
The granddaddy of spiders pushes back the evolutionary clock

How Sleep Cleans the Brain
A fresh look at your nightly brainwashing

Latest Stories
Can Plants Count?
It seems as though they can at least track the number of events in their environment

Making AI More Human
An interview with Berkeley researcher and author Nina Begus about her new book and proposal to fuse science and the humanities


From the newest issue
See more


Get the Nautilus newsletter
Cutting-edge science, unraveled by the brightest living thinkers.
Astronomy
See more AstronomyHubble Snaps a New Dazzling Photo of the Crab Nebula
It was formed by an explosion witnessed around the world almost a millennium ago

The Search for Alien Life Just Identified 45 New Targets
This subset of exoplanets are the most likely to be habitable


History
See more HistoryA Very Unscientific History of Scientific Hoaxes
The past, present, and future of academic deception
The Martyrs, Hunters, and Nature Lovers Who Came Together to Save Birds
An interview with James McCommons, author of The Feather Wars, about the past and future of bird conservation

How Three Students Designed an Atomic Bomb
A top-secret 1960s project tasked physics postdocs with building The Bomb

Psychology
See more PsychologyThe Students Who Believe Practice Makes Perfect Get Pretty Perfect Grades
There’s a reason it’s a popular aphorism


The Internet Has Not Killed Reading—or Attention Spans
An interview with Kevin Ashton, MIT technology pioneer and author of The Story of Stories

Get unlimited, ad-free Nautilus. Become a member today.
Environment
See more EnvironmentOrdinary Lab Gloves May Have Skewed Microplastic Data
That doesn’t mean microplastics aren’t a problem, though


These Seals Brave Polar Bear Country to Access an Ocean Buffet
Conservation plans for climate change must consider both fear and food

Zoology
See more Zoology

The Mystery of the Legless Lizards of Taiwan
The secretive reptile has confounded researchers for decades

Philosophy
See more PhilosophyA Light in the Dark: Finding the Good in the Natural World
Is it absurd to think that science can inform our values?

How ‘Tiny Shortcuts’ Are Poisoning Science
Seemingly harmless data tweaks are undermining the integrity of the entire field. We must define the problem to prevent it

Doing Science and Philosophy On Drugs
Justin Smith-Ruiu takes a philosophical and first-person look at psychedelics

The Big-Game Elephants Neanderthals Hunted for Food
A prehistoric butcher bonanza uncovered on an ancient German lakeshore

Read more
See all postsWho Gets to Do Science?
An interview with a neuroscientist who spent the last decade tearing down the class, race, and language barriers that keep people like him out of research


Now We Know What the Insects of the Jurassic Period Sounded Like
Thanks to crickets and relatives playing their “washboards”

The Giant Sloths and Armadillos of Prehistoric Texas
Snorkeling scientists uncover a treasure trove of megafauna fossils in a flooded cave

New Ape Fossil Could Shift Our Evolutionary Origins Northward
Ancestor of humans and other great apes turns up in Egypt


How Science Fiction Can Save Us
Three scientists share their bold vision for turning stories into testable experiments

Space Screws Up Sperm’s Ability to Navigate Properly
And not because they refuse to ask for directions
