What’s the oldest known living thing, and how do we know? Why should we even want to know? The explanation is a history of curiosity and care. It’s about our long-term relationships—spiritual and scientific—with long-lived plants, as long as long can be. It’s all about trees.
A tree is a plant that people call a tree—a term of dignity, not botany.
Although people construct the meaning of “trees” and assign age value to the vascular plants they call “ancient trees,” people cannot themselves create life that grows in place for centuries. Exclusively, solar-powered organisms enact that miracle. Among plants, there are ephemerals, annuals, biennials, perennials—and, beyond them all, perdurables, thousand-year woody life-forms.
As a rule, gymnosperms (flowerless plants with naked seeds) grow slower and live longer than angiosperms (flowering plants with fruits). Gymnosperms include ginkgo (a genus of one), cycads, and every kind of conifer—including yews, pines, firs, spruces, cedars, redwoods, cypresses, podocarps, and araucarias. All these lineages began hundreds of millions of years before the divergence of angiosperms. In effect, the newer, faster competition forced slow growers to retreat to exposed sites and poor soils, adverse niches conducive to oldness. Five thousand years is the approximate limit for nonclonal living under adversity. In plants, the potential for extreme longevity seems to be an evolutionary holdover from the deep past. Only about 25 plant species can, without human assistance, produce organisms that live beyond one millennium, and they are mainly conifers of primeval lineage. The cypress family contains the most perdurables, followed by the pine family. Many relict conifers hang on in limited, vulnerable habitats. The ice ages didn’t help their cause. In general, neither did humans, with their technologies of fire, domestication, and metalworking. Of some 600 conifer species, roughly one-third are endangered, with many genera reduced to a single species.
Each branch is effectively its own tree with its own history.
A gymnosperm doesn’t so much live long as die longer—or, live longer through dying. The interior dead wood—the heartwood—performs vital functions, mechanically and structurally. In comparison, the thin living outer layer is open to the elements. If damaged by an extrinsic event such as fire or lightning, this periderm doesn’t heal or scar like animal skin. Instead, new cambium covers the injury, absorbing it as one more historical record alongside its growth rings. Thus, an ancient conifer is neither timeless nor deathless, but timeful and deathful. A few special conifers such as bristlecone pine can live through sequential, sectorial deaths—compartmentalizing their external afflictions, shutting down, section by section, producing fertile cones for an extra millennium with the sustenance of a solitary strip of bark. The final cambium has vitality like the first. Longevity doesn’t suppress fecundity. Unlike animals, plants don’t accumulate proteins that lead to degenerative diseases.
The strongest correlation with long life (elongated death) is chemical. Longevous conifers produce copious resins—volatile, aromatic hydrocarbons like terpenes—that inhibit fungal rot and insect predation. Chemically, bristlecone is off the charts. Its high-elevation habitat offers additional protection from enemies, competitors, and fire, given that they tolerate dryness and cold. In habitats with chronic stress, conifers grow slower and stockier. Slow woody growth generates more lignin, another organic polymer with defensive properties. Stress-tolerant plants prioritize stability over size. Their stuntedness is equal parts adaptation and tribulation.
If dwarfism provides one path to longevity, gigantism provides another. Megaconifers such as sequoia in California, alerce in Chile, and kauri in New Zealand are more likely to survive any discrete attack. In cycles of forest regeneration, they tend to be first and last. They grow quickly as seedlings, establishing soil space, then keep on growing vertically, claiming canopy space. Long life is necessary because opportunities to establish new populations—conditions following intense disturbances—occur infrequently. This strategy comes with one big downside: the risk of falling. Gravity can be as fatal as rot. Another trade-off, the burden of transporting water skyward, explains why the tallest conifers occur in humid temperate zones.
Regrowth is another pathway to oldness, an adaptation that appears in both gymnosperms and angiosperms. Certain single-boughed woody species—notably ginkgo, redwood, yew, olive—can recover from catastrophic damage, even the death of the bole. These trees never lose their ability to resprout and regenerate. At the organismal level, they do not senesce, meaning they don’t lose vitality with age. In theory, such a plant is internally capable of immortality, though some external force inevitably ends its life. With particular species and cultivars, humans can force rejuvenation through grafting, pollarding, or coppicing. Plants that normally die young may live long under horticultural care.
Lastly, some perdurables, mainly angiosperms, grow as clonal colonies with many trunks, invisibly connected; their longevity is contained in the root mass—and the genome—rather than in the cambium aboveground. The clonal age is estimated from the date of sexual germination of the common ancestor. For example, a colony of quaking aspen may live—or, keep on dying—for tens of thousands of years.
Even nonclonal trees are de facto colonies and superorganisms. Treehood is as multitudinous as treeness is reductive. The underworld plant—physiologically and philosophically, the most generative part—is a networked system of roots that interact with other plants through symbiotic associations with fungi. Meanwhile, aboveground, each branch is effectively its own tree with its own history, as recorded in its unique shape. Modular organisms allow for infinite variety through plasticity. After a sudden disturbance, a tree can alter its growth trajectory. On a longer timescale, a population of woody plants can shift from arborescence to shrubbiness (or vice versa) in response to incremental change.
Humans multiplied and deplenished the Earth.
Relatively rapid microevolution at the population level belies the slowness of macroevolution at the species level. To be a woody plant is to be changeful and changeless at once, in different dimensions. On a twin-boughed bristlecone, the two stems are separated by four millennia of somatic mutations, yet that divergence is subtle compared to the fast-evolving bacteria and fungi that live on those branches, changing over millions of generations, all the while trying and largely failing to eat pinewood that might as well be wood from the geologic past. A half billion years after the development of lignin—one of the key outcomes of evolutionary history—lignophytes retain advantages.
The price of longevity is immobility. At the organismal level, a plant cannot migrate like an animal. Its localism is total. Trees take what comes until something indomitable comes along. Extrinsic mortality may result from a distinct catastrophe, such as fire or gale, or multiple, cumulative stressors. There are limits beyond which even the most deeply rooted organisms can no longer function. Thresholds of water, salinity, and temperature are absolute thresholds.
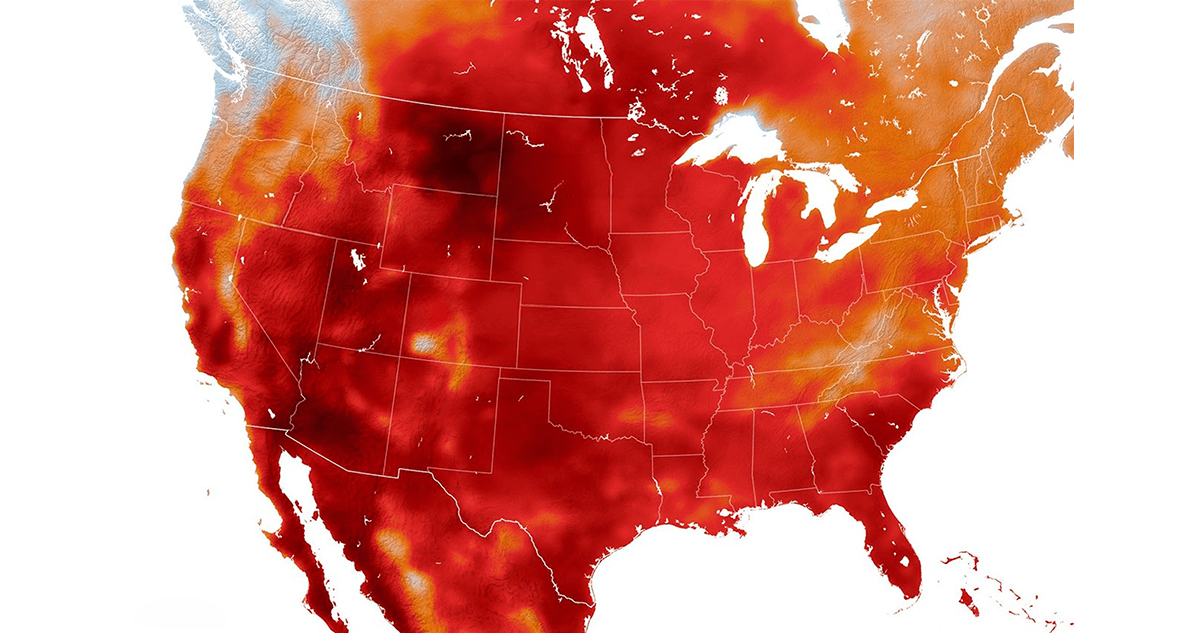
Climatic stability promotes arboreal longevity. The very recent geologic past was a favorable time for elderflora, big and small. According to computer models informed by tree-ring data, the immediate future will bring increased variability, extremity, and precarity. The famous “hockey stick” graph showing global mean temperature increase is one of many steep curves—population growth, resource use, methane and CO2 emissions, ocean acidification, and more—cumulatively known as the postwar “Great Acceleration.”
The flipside is the great diminution: fewer big trees and big animals; fewer old trees and old-growth forests; fewer old species and species overall. In biblical terms, humans multiplied and deplenished the Earth. Even protected areas are porous to pollution, desertification, illegal logging, invasive pests and pathogens, stronger storms, hotter droughts, rising temperatures, rising seas. Old Ones, wild and domesticated, face new vulnerabilities. In the United States West, past efforts to suppress wildfire have backfired spectacularly, priming forests for destructive megafires. Today’s elders of the land, monuments of lost climates, perish before their times—displaced by our times.
An ancient conifer is neither timeless nor deathless, but timeful and deathful.
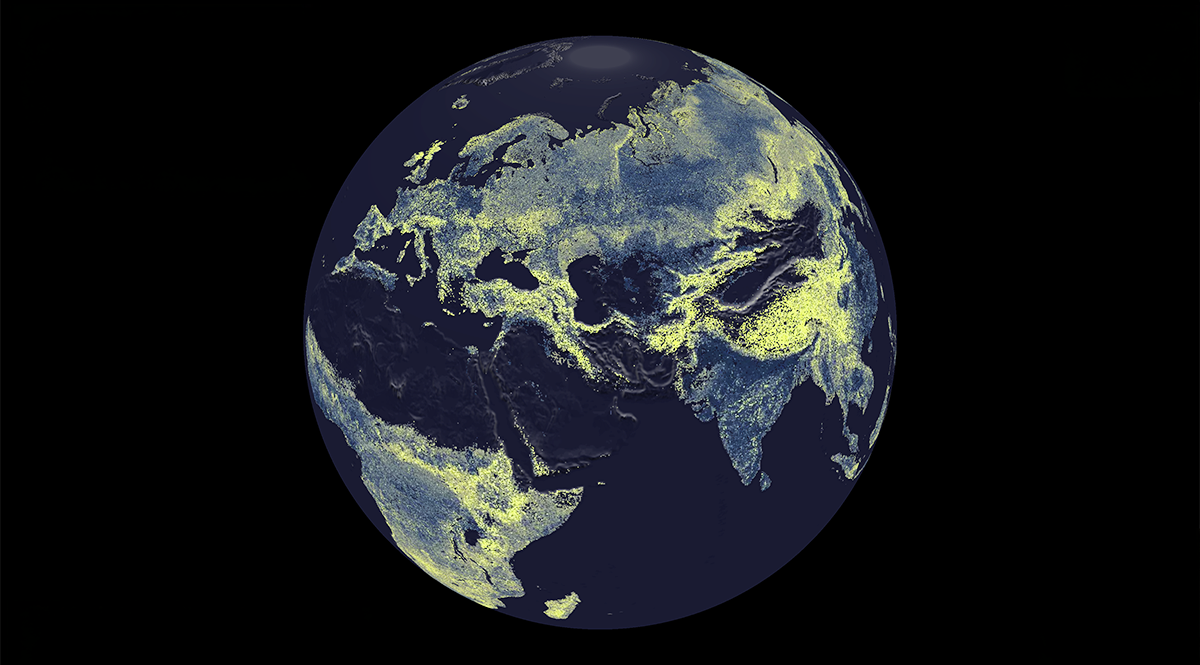
Forest dieback means a decline in chronodiversity. This concept relates to biodiversity, or the variety of life on Earth, crudely measured as the number of species, or “species richness.” Conservation biologists make a precautionary assertion about that enumeration. They argue that safeguarding the maximum possible amount of genetic information created over millions of years of evolutionary history is wise and moral. The complement of species richness is temporal richness. The biosphere has further possibilities if it contains species of various evolutionary ages, species of various life strategies and life spans, and specimens of various ages within species. It is an ecological loss of doubled magnitude when a species-rich, age-rich rain forest becomes row upon row of monocultural, monochronic crops. Over the past two centuries, states and corporations—often working against local users and Indigenous activists—have divided the forested areas of the globe into binary zones: large industrial plantations of “ordinary” young trees and small inviolate preserves of “extraordinary” old trees. Before the awareness of anthropogenic climate change, preservation through segregation had its own logic—the logic of permanence. National groves were supposed to last forever. Now, in a time of fateful dynamism, these outdoor museums of olden trees may be doomed by their fixity.
Does a naturally occurring tree of great age have value in itself? Foresters and forest ecologists have long debated this question. A century ago, technicians used words like “overage,” “overmature,” and “decadent” to describe standing timber past its prime. Commercial managers saw tree life as individual and rotational, and considered postmerchantable growth to be a biological waste of time. Their business—international markets for wood products—encouraged uniformity in age and size. By contrast, forest ecologists studied the communities in, on, and under each tree—each a world in itself—and saw forest life as processual. The cycle of life required dead and dying trees. Today, foresters meet ecologists halfway: Old trees provide nutrient cycling, carbon storage, and other “ecosystem services.”
Perdurables are so much more than service providers. They are gift givers. They invite us to be fully human—truly sapient—by engaging our deepest faculties: to venerate, to analyze, to meditate. They expand our moral and temporal imaginations.
In mythical form, trees appear in creation stories, present at time’s beginning. In graphical form, they represent seasons, cycles, genealogies, algorithms, and systems of knowledge. An olden bough is a bridge between temporalities we feel and those we can only think. This is why Darwin imagined millions of years of evolutionary history as a wide-spreading Tree of Life. Most profoundly, select living conifers—ancient organisms of ancient ancestry—are incarnations of geohistory. Volcanic eruptions, magnetic field reversals, and solar proton events leave signatures in their wood. Through tree-ring science, we see how woody plants register cyclical time and linear time, Chronos (durations) and Kairos (moments), climate and weather, the cosmogenic and the planetary. As multitemporal beings—short, long, and deep time together, in living form—perdurables allow us to think about the Anthropocene without anthropocentrism. They grant emotional access to timefulness. 
Jared Farmer is the Walter H. Annenberg Professor of History at the University of Pennsylvania, and the award-winning author of four books. He lives in Philadelphia.
Excerpted and lightly adapted from the book Elderflora by Jared Farmer. Copyright © 2022 by Jared Farmer. Reprinted by permission of Basic Books, an imprint of Perseus Books, LLC, a subsidiary of Hachette Book Group, Inc., New York, New York, USA. All rights reserved.
Lead image: Potapov Alexander / Shutterstock