



The first time I saw a free-living giraffe was in Tanzania’s Arusha National Park, where I was astounded by a yellow-and-brown head gliding gracefully and, it seemed, impossibly high above the tops of tall acacia trees. That was 11 years ago and underscored why the giraffe has remained one of my favorite animals. But it wasn’t just my African experiences that kept the giraffe in my graces.
As an evolutionary biologist and professor, I have put Giraffa camelopardalis on stage in my classrooms—well, not literally—as the embodiment of how natural selection has produced a creature that on the one hand is spectacularly adapted to its peculiar ecological niche and on the other is an example of evolution’s “clumsy, wasteful and blundering” process, to borrow Darwin’s own words. It’s sometimes assumed those blunders result from mutations or evolutionary errors. But in fact they result from history: the fact that at any given point natural selection has no choice but to work from what is already available. More than most animals, giraffes reflect the fact that organisms have not been created from scratch (or if they were, the Special Creator was notably inept). Rather, they have been cobbled together, via trial and success, from their historical antecedents.
Giraffes have long enjoyed a special place in the hearts of evolutionists for another reason: as a real-life example of how to distinguish Darwinian from Lamarckian evolution. It turns out, however, the story isn’t quite so simple; indeed, nothing about these fascinating and bizarre animals is. A close look at their private lives has implicated mating no less than munching when it comes to their notable anatomy, leading to issues that have spurred considerable debate among today’s (admittedly small) community of giraffologists.
Let’s start with the most obvious. The giraffe’s great height poses a number of difficulties that have been overcome via some notable adaptations. In order to pump blood seven feet above the animal’s heart to that towering head, an exceptionally high blood pressure is required, as much as three times the human systolic level. In order to keep from blowing their arteries apart, giraffes need special structural supports within those blood vessels.
Giraffes are an extraordinary example of evolution’s “clumsy, wasteful and blundering” process, to borrow Darwin’s own words.
In the other direction, to prevent blood from pooling in their feet, which are at the end of some very long legs, giraffes have evolved the equivalent of compression stockings, like those that people use post-surgery or to prevent deep vein thrombosis on long airplane rides.1 The giraffe’s hack consists of highly elastic blood vessel walls, combined with an extensive capillary bed. By restricting perfusion of fluid into surrounding tissues, these structures keep a giraffe’s blood in its vessels, where it belongs, rather than in surrounding tissue. These animals have another specially adapted compression system in their necks, which prevents too much blood from rushing to their heads when they bend down to drink—which they don’t do very often, since they get most of their water from the leaves they eat, using their highly flexible 18-inch tongue.
As notably long as are giraffe necks, these are actually outclassed by their legs, such that those monumental necks are—believe it or not—too short to comfortably reach a puddle; as a result, a drinking giraffe must splay its front feet wide apart. And, by the way, the same fluid sluice-way control mechanism in its lengthy neck works in reverse when a giraffe is done drinking and eventually raises its high head, allowing only a relative trickle of blood to flow back down so that its brain doesn’t suddenly become hypoxic.2
The name “giraffe” comes from the Arabic word “zarafah,” which means “one who walks swiftly,” and giraffes use those long legs to walk swiftly indeed, although given their long leggy levers and somewhat swaying, vertically balanced pillar of a neck, they actually appear to move slowly. For reasons that are unclear, but presumably are the result of genuinely giraffish biomechanics, when giraffes have occasion to move swiftly, they don’t canter or gallop like nearly all other quadrupeds, which do so by alternating front and rear legs off the ground. Rather, they move front and rear legs on the same side at the same time, alternating left and right—rather than front and back—in a pattern known as “pacing.” It’s actually quite efficient and graceful, more gainly than ungainly, once the human observer gets over its novelty. (Incidentally, camels also pace,3 which might explain the Latin species name for giraffes: camelopardali, because of the ancient belief that giraffes were part camel and part leopard, the latter because of giraffes’ notably blotchy, flagstone-looking camouflage.)
Then there’s that notable neck. How many bones comprise a giraffe’s neck? Answer: seven (a 1999 Journal of Zoology study says eight),4 the same number as in a human being, except that the giraffe’s are huge, each cervical vertebra averaging about 10 inches in length. And in a mouse? Also seven, although—surprise—each is rather small. Mouse-like, in fact.
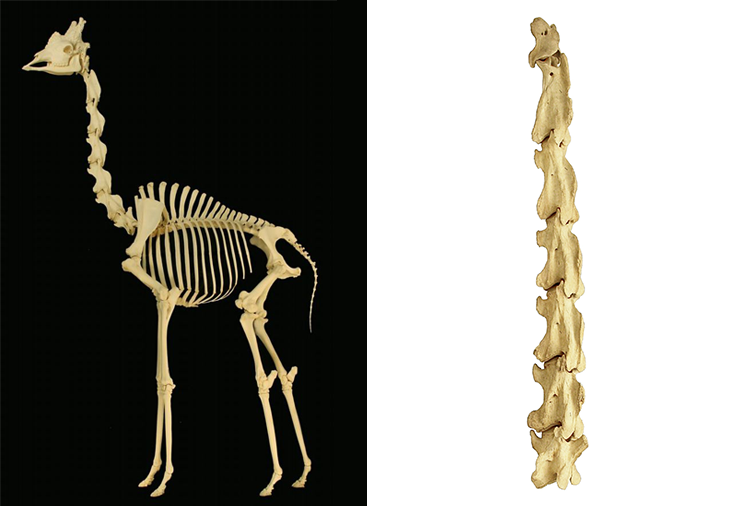
Probably the most famous thing about the giraffe’s neck (at least among biologists), is a peculiarity of its nerves, notably the left recurrent laryngeal nerve, which sends impulses to the animal’s larynx. Forget for the moment that giraffes are probably the quietest of any large mammal; they do vocalize a little, albeit faintly. No one knows why they are so soft-spoken, although it is clear that their larynx gets the same basic innervation as other, noisier mammals, albeit after the impulse has traveled farther. Perhaps it has something to do with that long neck, or maybe they just don’t have much to say. In any event, the muscles of a giraffe’s larynx—as with ours—are stimulated by their laryngeal nerve, and this is where we encounter one of zoology’s most stunning examples of extraordinarily unintelligent design.
The laryngeal nerves, present in vertebrates generally, branch off from the larger vagus nerve, the major contributor to every vertebrate’s parasympathetic system, which sends signals to all of the major internal organs, including the heart and digestive system. In all mammals, the recurrent laryngeal nerves depart from the vagus at the level of the aortic arch, just where the aorta—initially ascending from the heart and then sending the carotid arteries to nourish the head and neck—dives posteriorly to provide blood to the rest of the body. This arch of the aorta makes a hairpin 180-degree loop, which is no problem for the right recurrent laryngeal nerve, which, being on the “correct” side, avoids the aortic bend and goes directly up to the larynx, along the trachea. Its left counterpart, on the other hand, is forced to curve under the aortic arch before heading larynx-ward: a bit anatomically inconvenient but not a major problem in most vertebrates, including human beings, since this loopy path only necessitates a few extra inches of length.
Herein lies both an interesting dilemma for long-necked creatures as well as an object lesson in evolution. Among modern fish, and presumably the fishy ancestors of today’s mammals, the recurrent laryngeal nerves (left and right) would have followed a straight path from the brain, along the heart and then to the gills. It’s pretty much the same in short-necked early mammals, although the left branch, stuck under the downward curving side of the aortic arch, would have had a slightly longer, loopier route. But among those critters that evolved long necks, the poor left recurrent laryngeal nerve was forced to perform a downright ridiculous detour during embryonic development: emerging from the brain, going southward so as to loop just below that ever-retreating aortic arch, then heading upward again, eventually to reach the larynx.
The “necks for sex” hypothesis states long necks were selected because they contributed to success in male-male competition.
In the case of giraffes, this absurd arrangement now necessitates a nerve that’s about 15 feet long (7.5 feet down and then back up), whereas if it had simply been routed directly, its entire length would have been perhaps six inches. Why this weirdness? Because the evolution of giraffes proceeded from the immediate antecedents of giraffes, whose evolution proceeded from theirs in turn, going back to a common ancestral fish, whose left recurrent laryngeal nerves were perfectly reasonable, thank you. (And by the way, don’t expend all your recurrent laryngeal sympathy to giraffes: There were other vertebrate descendants of fish—notably the sauropod dinosaurs—whose 45-foot-long necks would have necessitated a lot more nerve: roughly 90 feet! A recent technical account—in the journal Acta Palaeontological Polinica, to be exact—described this absurdity as a “monument of inefficiency.”5)
So far, so awkward. But why have modern giraffes evolved such troublesome long necks in the first place? Here’s where giraffes help us distinguish Darwinian from Lamarckian evolution. The Lamarckian take is giraffe necks became long because their ancestors kept stretching theirs. The Darwinian view is early giraffes differed with respect to their neck length, and those with longer ones reproduced more successfully. According to Lamarck’s notion of the “inheritance of acquired characteristics”—not that different from “use ’em or lose ’em”— organs enlarge when used and contract when ignored. Pretty much everyone agrees ancestral giraffes started with necks not unlike other ungulates.
But Lamarck and his followers imagined that as ancient giraffes stretched to reach leaves higher and higher at the tops of savannah trees (and giraffes are indeed browsers), their necks elongated, just as muscles get bigger with use, until over time, their descendants had the longest necks around. The Darwinian perspective is subtly different, and more accurate: As competition for leafy vegetation developed, those ancestral giraffes that had somewhat longer necks were just a teeny bit more able to nourish themselves by reaching high-growing leaves.
So they left more offspring, who in turn had longer necks, like their parents. Natural selection thus favored long necks in this lineage, along with the necessary adaptations such as the ability to generate and withstand ultra-high blood pressure while preventing blood from pooling at the feet, plus the unavoidable downsides of those deviously routed left recurrent laryngeal nerves.
But the story isn’t over. There is another, competing explanation for why giraffes have such long necks, this one involving sexual selection. First, a quick detour. Although many people assume otherwise, there is no fundamental difference between natural selection and sexual selection. Natural selection occurs any time differential reproduction is afoot, with some genes more successful than others in projecting copies of themselves into future generations. Sexual selection involves competition to acquire and obtain a mate, and is every bit as natural as other forms of selection called upon when it comes to, say, obtaining food, avoiding predators, sleeping when tired, or scratching when itchy. It’s just that sexual selection sometimes results in traits—the classic one being the peacock’s tail—that are disadvantageous in the ecological realm, but nonetheless positively selected because on balance they contribute to fitness, via same-sex competition or opposite sex attraction, more than they cost in the straightforward realm of pure survival.
It turns out that male giraffes are less cuddly and more ornery than they appear. During the rut, they go at each other, using their heavy, bony heads at the end of their long flexible necks roughly like a medieval ball-and-chain weapon, or flail. And the longer the neck, the more force behind each blow. The prospect—colorfully known as the “necks for sex” hypothesis—arises that long necks were positively selected because they contributed to success in male-male competition. Also, females may have preferred studly, long-necked males, at least in part because their offspring would likely have long necks. That would help males in battles with other males and make them sexually attractive to the next generation of choosing females. Either way, long-neck genes in males, and genes in females that influenced their preference for long-neck males, would be passed along.6 This is a variant of what behavioral ecologists call the “sexy son hypothesis,” applied as well to the evolution of peacocks’ tails and other seemingly outlandish male traits. According to this hypothesis, female giraffes evolved their long necks largely as a by-product of sexual selection among males.
The idea is controversial, and the final word is not yet in.7 Favoring the “necks for sex” hypothesis is the finding that males with longer necks really are more successful in male-male combat and, moreover, females really do prefer elongated necks in their potential mates (which gives new dimension to the term “necking”). Also, most giraffe foraging takes place at shoulder level rather than when the animals have their necks maximally extended. In addition, giraffes are fully six feet taller than other competing browsers, which would seem to argue that competitive foraging as such hasn’t been the main driving pressure behind their altitudinal evolution. On the other hand, maybe it’s during times of food shortage that being extra tall pays off, and, moreover, plants tend to concentrate the most nutritious part of their foliage in the top-most, growing tips.8
There is no reason, however, to think that foraging success and sexual success are mutually exclusive in their possible evolutionary impact.9 That prospect is another example of how these lanky, leggy, long-necked giants embody the diversity of life sculpted by evolution. At the same time, it can be challenging to imagine a creature as improbable as a giraffe engaging in sex at all, not to mention having been shaped by its exigencies. But the same thing applies to contemplating sex among our own parents, and yet the fact that we are here bespeaks that somehow they—like giraffes—managed it, at least upon occasion.
David P. Barash is an evolutionary biologist and professor of psychology at the University of Washington. His most recent book is Buddhist Biology: Ancient Eastern Wisdom Meets Modern Western Science.
References
1. Hargens, A.R., Millard, R.W., Pettersson, K., & Johansen, K. Gravitational haemodynamics and oedema prevention in the giraffe. Nature 329, 59-60 (1987).
2. Mitchell, G. & Skinner, J.D. How giraffes adapt to their extraordinary shape. Transactions of the Royal Society of South Africa 48, 207-218 (1993).
3. Dagg, A.I. The locomotion of the camel (Camelus dromedarius). Journal of Zoology 174, 67-78 (1974).
4. Solounias, N. The remarkable anatomy of the giraffe’s neck. Journal of Zoology 247, 257-268 (1999).
5. Wedel, M.J. A monument of inefficiency: The presumed course of the recurrent laryngeal nerve in sauropod dinosaurs. Acta Palaeontologica Polonica 57, 251-256 (2011).
6. Simmons, R.E. & Scheepers, L. Winning by a neck: Sexual selection in the evolution of giraffe. American Naturalist 148, 771-786 (1996).
7. Mitchell, G., Van Sittert, S.J., & Skinner, J.D. Sexual selection is not the origin of long necks in giraffes. Journal of Zoology 278, 281-286 (2009).
8. Cameron, E.Z. & du Toit, J.T. Winning by a neck: Tall giraffes avoid competing with shorter browsers. The American Naturalist 169, 130-135 (2007).
9. Simmons, R.E. & Altwegg, R. Necks‐for‐sex or competing browsers? A critique of ideas on the evolution of giraffe. Journal of Zoology 282, 6-12 (2010).
Lead photo © vince42/Flickr









